Anpassungsstrategien
Uns interessiert, welche physiologischen, biochemischen und molekularen Anpassungen haben Algen und Cyanobakterien entwickelt, um extreme Lebensräume zu erobern. Lebensräume, in denen die Mehrheit der Lebewesen nicht lebensfähig ist. Die Anpassungsmechanismen sind aus Sicht der Grundlagenforschung und der Biotechnologie relevant: Sie helfen, die Lebensvorgänge unter Extrembedingungen zu verstehen und die "Erfindungen der Natur" für den Menschen nutzbar zu machen.
Und wir wollen wissen, ob und in welcher Weise die Organismen auf die Veränderungen der aquatischen und terrestrischen Lebensräume reagieren, die der Klimawandel mit sich bringt.
Unseren Fokus richten wir dabei auf Anpassungen an:
Extreme Temperaturen
Aeroterrestrische Algen auf Gebäudeoberflächen oder in hochalpinen Bodenkrusten sind extremen Temperaturschwankungen ausgesetzt. Ein Wetterwechsel kann eine Änderung der Maximal- oder Minimaltemperatur von bis zu 15 Grad Celsius mitbringen. Darüber hinaus betragen die Temperaturunterschiede zwischen Tag und Nacht in unseren Breiten bis zu 15 Grad Celsius. An Sommertagen schwankt die Tag- und Nachttemperatur auf Dachziegeln mehr als 50 Grad. An diesen Standorten siedelnde Organismen müssen temperaturresistent sein, um dort zu überleben.
Daher untersuchen wir das Wachstum und die Photosynthese unter verschiedenen Temperaturen und ermitteln Optima sowie obere und untere Grenzwerte. Auf die Messdaten wenden wir mathematische Modelle an.
Dagegen sind arktische Kieselalgen auf dem Meeresboden ganzjährig mit sehr kalten Temperaturen konfrontiert. Uns interessiert, welche ökologische und ökophysiologische Leistungsfähigkeit diese phototrophen Biofilme im Eismeer haben.
PAR- und UV-Belastung
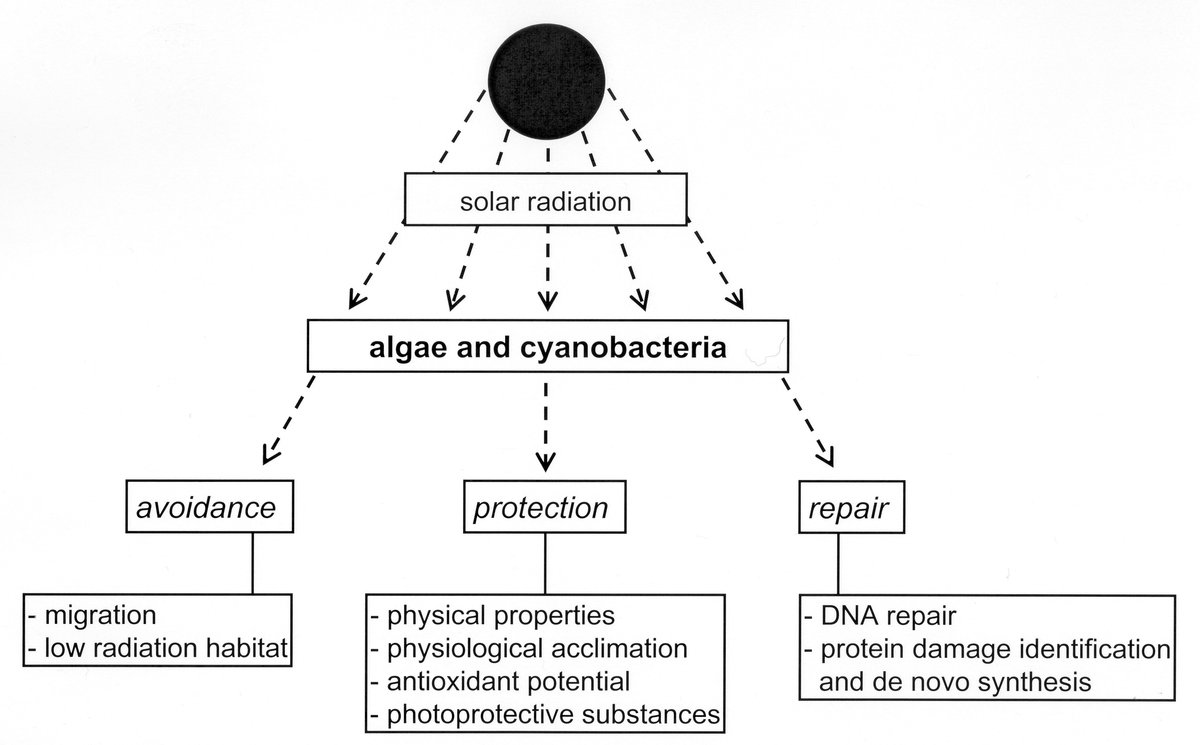
Viele phototrophe Organismen stecken in einem Dilemma: Auf der einen Seite benötigen sie ausreichend Sonnenstrahlung für die Photosynthese (photosynthetisch aktive Strahlung, PAR: 400-700 nm), auf der anderen Seite kämpfen sie im Sommer mit einem Überangebot, was die Photosynthese hemmen und den Photosyntheseapparat schädigen kann. Deshalb reduzieren sie unter hohen Lichtintensitäten empfindliche Komponenten des Photosynthese-Apparates und aktivieren Schutzmechanismen. Manche bilden Schutzpigmente, wie beispielsweise ß-Carotin, die überschüssige Strahlungsenergie als Wärme abgegeben. Und sie müssen die zerstörenden Oxidationsvorgänge unterdrücken, die mit Starklicht einhergehend. Dazu bilden sie verschiedene Antioxidantien und aktivieren antioxidative Enzyme.
Ein weiteres ökologisches Problem ist die Erhöhung der harten ultravioletten Strahlung (UV-B) in Mitteleuropa. UV-B (Wellenlänge: 280-315 nm) wirkt stark erbgutverändernd und schädigt zahlreiche Biomoleküle in der Zelle, wodurch viele Lebensäußerungen und Leistungsfähigkeiten der Algen negativ beeinflusst werden. Viele Algen und Cyanobakterien schützen sich daher mit spezifischen Substanzen, die als Sonnenschutz wirken.
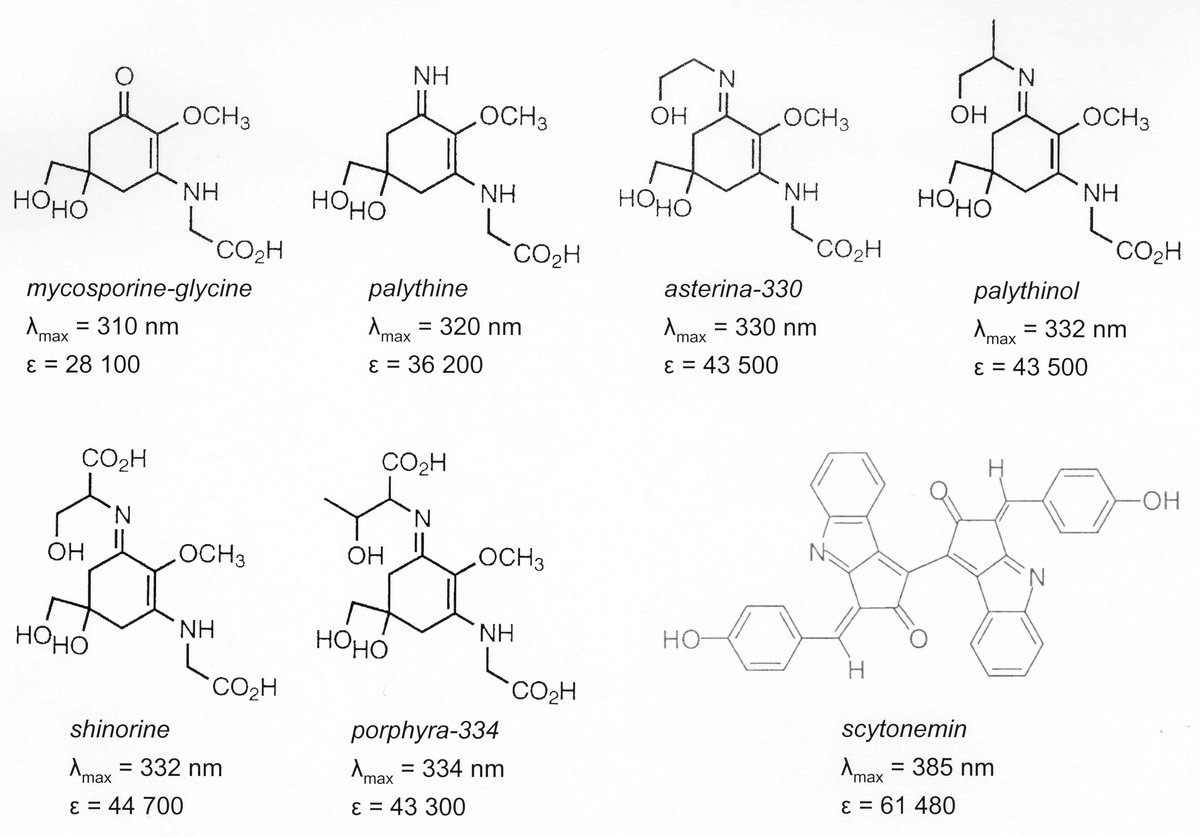
Die Fähigkeit zur Bildung UV-protektiver mykosporin-ähnlicher Aminosäuren (MAAs) konnte bei Flachwasser-Rotalgen und aeroterrestrischen Grünalgen nachgewiesen werden. Bisher sind etwa 25 verschiedene MAAs aus unterschiedlichen Organismen bekannt. Im Zentrum unserer Forschung steht der MAA-Metabolismus und wie er durch abiotische und biotische Faktoren beeinflusst wird. Wir führen unsere Analysen vorwiegend unter kontrollierten Bedingungen mit Laborkulturen durch.
Trockenheit

Aeroterrestrische Algen auf Gebäudeoberflächen und in Bodenkrusten sind häufig Trockenstress ausgesetzt. Nach Niederschlägen ist reichlich Wasser vorhanden, fehlt aber im Sommer dagegen ganz. Diese Mikroalgen bilden extrem dicke Zellwände oder Schleimhüllen, die als Verdunstungsschutz dienen. In lang anhaltenden Trockenzeiten kapseln sie Sporen ab, die dicke, imprägnierte Zellwände haben und ein jahrelanges Überleben in Trockenheit garantieren. Kommen die Sporen wieder mit Wasser in Kontakt, keimen sie nach kürzester Zeit wieder aus. Auch zellbiologische Komponenten - wie das Zytoskelett - stabilisieren die Struktur der Algenzelle bei Trockenstress.
Wie unsere ökophysiologischen Untersuchungen zeigen, wachsen aeroterrestrischer Grünalgen, auch ohne flüssiges Wasser. Ihnen reichen 100 Prozent relative Luftfeuchte zum Wachsen und Photosynthesetreiben. Sie sind darüber hinaus fähig, ungewöhnliche organische Substanzen zu synthetisieren und in hoher Konzentration anzusammeln. Insbesondere Zuckeralkohole erfüllen gleich mehrere verschiedene Stoffwechselfunktionen unter ungünstigen Umweltbedingungen: Sie wirken der drohenden „Aufkonzentrierung“ von Salzen unter Trockenstress entgegen, stabilisieren als Thermoschutz empfindliche Proteine und liefern Energie für den Überlebensstoffwechsel sowie für Reparaturprozesse.
Salz-Stress


Auf den Luftwurzeln der Mangrovenpflanzen siedeln einzigartige Rotalgen, die ökologisch und bezüglich ihrer Inhaltsstoffe eine außergewöhnliche Gruppe darstellen. In den Mangroven wechselt mit den Gezeiten ständig der Salzgehalt des Mediums. Diese Pflanzen sind jedoch in der Lage eine aktive Akklimation durchzuführen. Dazu regulieren sie den intrazellulären Gehalt osmotisch aktiver Substanzen.
Während die meisten Rotalgen als Hauptphotosyntheseprodukt und organisches Osmolyt das so genannte Heterosid Floridosid (α-D-Galaktopyranosyl-(1-2)-Glyerol) bilden, weisen Mangrovenalgen stattdessen die Zuckeralkohole Mannitol, Dulcitol und Sorbitol auf. Mannitol und andere Zuckeralkohole werden unter Salz-Stress von den Zellen akkumuliert, und üben neben ihrer Funktion als organische Osmolyte, eine wichtige Rolle als so genannte compatible solutes aus. Compatible solutes stablisieren - im Gegensatz zu anorganischen Osmolyten (wie Na+ und Cl-) - Enzyme, Membranen und Strukturmakromoleküle.
Auf den Luftwurzeln der Mangrovenpflanzen siedeln einzigartige Rotalgen, die ökologisch und bezüglich ihrer Inhaltsstoffe eine außergewöhnliche Gruppe darstellen. In den Mangroven wechselt mit den Gezeiten ständig der Salzgehalt des Mediums. Diese Pflanzen sind jedoch in der Lage eine aktive Akklimation durchzuführen. Dazu regulieren sie den intrazellulären Gehalt osmotisch aktiver Substanzen.
Während die meisten Rotalgen als Hauptphotosyntheseprodukt und organisches Osmolyt das so genannte Heterosid Floridosid (α-D-Galaktopyranosyl-(1-2)-Glyerol) bilden, weisen Mangrovenalgen stattdessen die Zuckeralkohole Mannitol, Dulcitol und Sorbitol auf. Mannitol und andere Zuckeralkohole werden unter Salz-Stress von den Zellen akkumuliert, und üben neben ihrer Funktion als organische Osmolyte, eine wichtige Rolle als so genannte compatible solutes aus. Compatible solutes stabilisieren - im Gegensatz zu anorganischen Osmolyten (wie Na+ und Cl-) - Enzyme, Membranen und Strukturmakromoleküle.
Nach gegenwärtigem Kenntnisstand ist innerhalb der roten Makroalgen Caloglossa die einzige Gattung, die einen Mannitol-Metabolismus aufweist. Ausgehend von Fruktose-6-Phosphat wird Mannitol im Zytoplasma über die Schlüsselenzyme Mannitol-1-Phosphat Dehydrogenase und Mannitol-1-Phosphatase synthetisiert. Der Mannitol-Abbau zu Fruktose-6-Phosphat wird in Caloglossa von einer Mannitol Dehydrogenase und einer Hexokinase katalysiert. Der vollständige, d.h. anabolische und katabolische, Stoffwechselweg des Mannitols ist als so genannter Mannitol-Zyklus bisher nur in einigen Pilzen, parasitischen Protozoen und Caloglossa leprieurii sowie kürzlich in der primitiven einzelligen Rotalge Dixoniella grisea beschrieben.
Kohlendioxid-Erhöhung
Kohlendioxid lässt nicht nur die Temperaturen in der Atmosphäre steigen und verursacht damit den globalen Klimawandel, sondern lässt auch die Ozeane saurer werden. Die zunehmende Versauerung der Ozeane ist inzwischen zweifelsfrei belegt. Die rasante Geschwindigkeit, mit der der Säuregrad des Meerwassers steigt, ist dabei beispiellos in den vergangenen 20 Millionen Jahren. Als Folge der Ozeanversauerung verändert sich auch die Chemie des Meerwassers, was sich vor allem in einer deutlichen Abnahme der Karbonat-Ionen widerspiegelt. Karbonat dient als Baustein für alle Kalk bildenden Tiere und Pflanzen der Ozeane. Ohne Karbonate fehlt etwa Muscheln und Schnecken der notwendige Baustoff für den Schalenbau.
Welche weit reichenden Konsequenzen diese Prozesse für die Ökosysteme in den Ozeanen und damit auch für den Menschen bergen, ist allerdings noch weitgehend unklar. Klar ist lediglich, dass von der Fischerei bis hin zum Tourismus alle Wirtschaftsbranchen betroffen sein werden, die ihren Lebensunterhalt aus dem Meer erzielen.
Zur Übersicht unserer aktuellen Projekte